
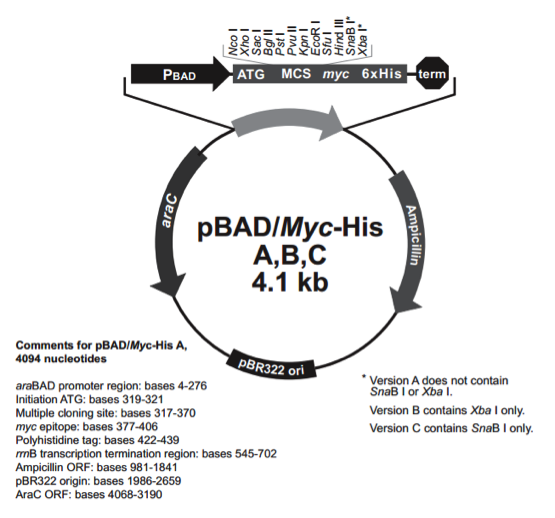
pBad/Myc-His C
pBad/Myc-His C
编号 | 载体名称 |
北京华越洋生物VECT5110 | pBad/Myc-His C |
pBadMyc-His C载体基本信息
载体名称: | pBAD/Myc-His C |
质粒类型: | 大肠杆菌表达载体;诱导表达载体 |
高拷贝/低拷贝: | 低拷贝 |
克隆方法: | 限制性内切酶;多克隆位点 |
启动子: | araBAD |
载体大小: | 4093 bp |
5' 测序引物及序列: | pBAD Forward: 5′-ATGCCATAGCATTTTTATCC-3′ |
3' 测序引物及序列: | pBAD Reverse 5′-GATTTAATCTGTATCAGG-3′ |
载体标签: | 6x His Tag(C-端),c-Myc Epitope(C-端) |
载体抗性: | 氨苄青霉素(Ampicillin) |
克隆菌株: | TOP10 |
表达菌株: | 推荐LMG194 |
备注: | pBAD/Myc-His C载体是阿拉伯糖调控载体; |
稳定性: | 稳表达 |
组成型/诱导型: | 诱导型(阿拉伯糖) |
病毒/非病毒: | 非病毒 |
pBadMyc-His C载体质粒图谱和多克隆位点信息
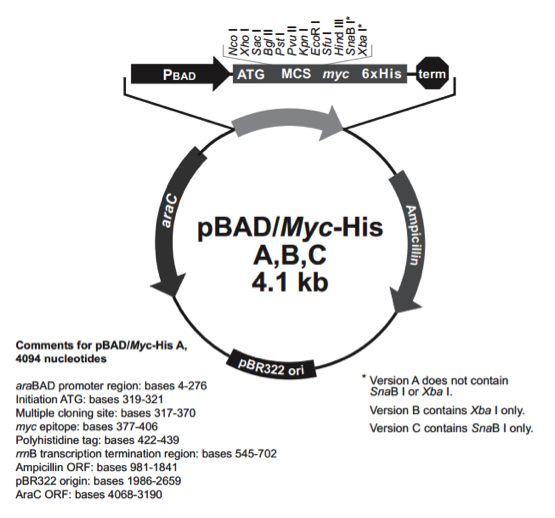

pBadMyc-His C载体简介
pBAD/His和PBAD/Myc-His载体质粒是衍生于pBR322载体。载体设计用来在大肠杆菌中进行可调节,剂量依赖性的表达和纯化重组目的蛋白。使用大肠杆菌araBAD启动子(pBAD)增强了大肠杆菌重组蛋白可溶性表达的水平。pBAD/His和pBAD/Myc His载体上的调节蛋白AraC能够调控pBad启动子。
pBAD/Myc-His A,B,C 载体简介
The pBAD/His and pBAD/Myc-His plasmids are pBR322-derived expression vectors designed for regulated, dose-dependent recombinant protein expression and purification in E. coli. Optimum levels of soluble, recombinant protein are possible using the araBAD promoter (PBAD) from E. coli. The regulatory protein, AraC, is provided on the pBAD/His and pBAD/Myc-His vectors allowing regulation of PBAD.
The pBAD/Myc-His Kit provides all of the necessary reagents to express your protein in a tightly regulated fashion. The pBAD/Myc-His vector expresses native proteins or fusion proteins with a C-terminal tag. The vector provides:
Þ The araBAD promoter for tightly regulated expression
Þ Translation initiation signals optimized for E. coliexpression
Þ C-terminal polyhistidine (6xHis) tag for purification with nickel-chelating resin or detection with an Anti-His(C-term) Antibody
Þ C-terminal c-myc epitope for detection and analysis with an Anti-myc Antibody
Three vectors are provided (A, B, and C). Each has the C-terminal tag in a different reading frame relative to the multiple cloning site to simplify in-frame cloning of your gene.
L-阿拉伯糖调控表达
In the presence of L-arabinose, expression from PBAD is turned on while the absence of L-arabinose produces very low levels of transcription from PBAD (Lee, 1980; Lee et al., 1987). Uninduced levels are repressed even further by growth in the presence of glucose. Glucose reduces the levels of 3′,5′-cyclic AMP, thus lowering expression of the catabolite-repressed PBAD promoter (Miyada et al., 1984). By varying the concentration of L-arabinose, protein expression levels can be optimized to ensure maximum expression of soluble protein. In addition, the tight regulation of PBAD by AraC is useful for expression of potentially toxic or essential genes (Carson et al., 1991; Dalbey and Wickner, 1985; Guzman et al., 1992; Kuhn and Wickner, 1985; Russell et al., 1989; San Millan et al., 1989). For more information on the mechanism of expression and repression of the ara regulon, refer to Schleif, 1992.
pBadMyc-His C载体序列
ORIGIN
1 AAGAAACCAA TTGTCCATAT TGCATCAGAC ATTGCCGTCA CTGCGTCTTT TACTGGCTCT
61 TCTCGCTAAC CAAACCGGTA ACCCCGCTTA TTAAAAGCAT TCTGTAACAA AGCGGGACCA
121 AAGCCATGAC AAAAACGCGT AACAAAAGTG TCTATAATCA CGGCAGAAAA GTCCACATTG
181 ATTATTTGCA CGGCGTCACA CTTTGCTATG CCATAGCATT TTTATCCATA AGATTAGCGG
241 ATCCTACCTG ACGCTTTTTA TCGCAACTCT CTACTGTTTC TCCATACCCG TTTTTTGGGC
301 TAACAGGAGG AATTAACCAT GGATCCGAGC TCGAGATCTG CAGCTGGTAC CATATGGGAA
361 TTCGAAGCTT ACGTAGAACA AAAACTCATC TCAGAAGAGG ATCTGAATAG CGCCGTCGAC
421 CATCATCATC ATCATCATTG AGTTTAAACG GTCTCCAGCT TGGCTGTTTT GGCGGATGAG
481 AGAAGATTTT CAGCCTGATA CAGATTAAAT CAGAACGCAG AAGCGGTCTG ATAAAACAGA
541 ATTTGCCTGG CGGCAGTAGC GCGGTGGTCC CACCTGACCC CATGCCGAAC TCAGAAGTGA
601 AACGCCGTAG CGCCGATGGT AGTGTGGGGT CTCCCCATGC GAGAGTAGGG AACTGCCAGG
661 CATCAAATAA AACGAAAGGC TCAGTCGAAA GACTGGGCCT TTCGTTTTAT CTGTTGTTTG
721 TCGGTGAACG CTCTCCTGAG TAGGACAAAT CCGCCGGGAG CGGATTTGAA CGTTGCGAAG
781 CAACGGCCCG GAGGGTGGCG GGCAGGACGC CCGCCATAAA CTGCCAGGCA TCAAATTAAG
841 CAGAAGGCCA TCCTGACGGA TGGCCTTTTT GCGTTTCTAC AAACTCTTTT GTTTATTTTT
901 CTAAATACAT TCAAATATGT ATCCGCTCAT GAGACAATAA CCCTGATAAA TGCTTCAATA
961 ATATTGAAAA AGGAAGAGTA TGAGTATTCA ACATTTCCGT GTCGCCCTTA TTCCCTTTTT
1021 TGCGGCATTT TGCCTTCCTG TTTTTGCTCA CCCAGAAACG CTGGTGAAAG TAAAAGATGC
1081 TGAAGATCAG TTGGGTGCAC GAGTGGGTTA CATCGAACTG GATCTCAACA GCGGTAAGAT
1141 CCTTGAGAGT TTTCGCCCCG AAGAACGTTT TCCAATGATG AGCACTTTTA AAGTTCTGCT
1201 ATGTGGCGCG GTATTATCCC GTGTTGACGC CGGGCAAGAG CAACTCGGTC GCCGCATACA
1261 CTATTCTCAG AATGACTTGG TTGAGTACTC ACCAGTCACA GAAAAGCATC TTACGGATGG
1321 CATGACAGTA AGAGAATTAT GCAGTGCTGC CATAACCATG AGTGATAACA CTGCGGCCAA
1381 CTTACTTCTG ACAACGATCG GAGGACCGAA GGAGCTAACC GCTTTTTTGC ACAACATGGG
1441 GGATCATGTA ACTCGCCTTG ATCGTTGGGA ACCGGAGCTG AATGAAGCCA TACCAAACGA
1501 CGAGCGTGAC ACCACGATGC CTGTAGCAAT GGCAACAACG TTGCGCAAAC TATTAACTGG
1561 CGAACTACTT ACTCTAGCTT CCCGGCAACA ATTAATAGAC TGGATGGAGG CGGATAAAGT
1621 TGCAGGACCA CTTCTGCGCT CGGCCCTTCC GGCTGGCTGG TTTATTGCTG ATAAATCTGG
1681 AGCCGGTGAG CGTGGGTCTC GCGGTATCAT TGCAGCACTG GGGCCAGATG GTAAGCCCTC
1741 CCGTATCGTA GTTATCTACA CGACGGGGAG TCAGGCAACT ATGGATGAAC GAAATAGACA
1801 GATCGCTGAG ATAGGTGCCT CACTGATTAA GCATTGGTAA CTGTCAGACC AAGTTTACTC
1861 ATATATACTT TAGATTGATT TAAAACTTCA TTTTTAATTT AAAAGGATCT AGGTGAAGAT
1921 CCTTTTTGAT AATCTCATGA CCAAAATCCC TTAACGTGAG TTTTCGTTCC ACTGAGCGTC
1981 AGACCCCGTA GAAAAGATCA AAGGATCTTC TTGAGATCCT TTTTTTCTGC GCGTAATCTG
2041 CTGCTTGCAA ACAAAAAAAC CACCGCTACC AGCGGTGGTT TGTTTGCCGG ATCAAGAGCT
2101 ACCAACTCTT TTTCCGAAGG TAACTGGCTT CAGCAGAGCG CAGATACCAA ATACTGTCCT
2161 TCTAGTGTAG CCGTAGTTAG GCCACCACTT CAAGAACTCT GTAGCACCGC CTACATACCT
2221 CGCTCTGCTA ATCCTGTTAC CAGTGGCTGC TGCCAGTGGC GATAAGTCGT GTCTTACCGG
2281 GTTGGACTCA AGACGATAGT TACCGGATAA GGCGCAGCGG TCGGGCTGAA CGGGGGGTTC
2341 GTGCACACAG CCCAGCTTGG AGCGAACGAC CTACACCGAA CTGAGATACC TACAGCGTGA
2401 GCTATGAGAA AGCGCCACGC TTCCCGAAGG GAGAAAGGCG GACAGGTATC CGGTAAGCGG
2461 CAGGGTCGGA ACAGGAGAGC GCACGAGGGA GCTTCCAGGG GGAAACGCCT GGTATCTTTA
2521 TAGTCCTGTC GGGTTTCGCC ACCTCTGACT TGAGCGTCGA TTTTTGTGAT GCTCGTCAGG
2581 GGGGCGGAGC CTATGGAAAA ACGCCAGCAA CGCGGCCTTT TTACGGTTCC TGGCCTTTTG
2641 CTGGCCTTTT GCTCACATGT TCTTTCCTGC GTTATCCCCT GATTCTGTGG ATAACCGTAT
2701 TACCGCCTTT GAGTGAGCTG ATACCGCTCG CCGCAGCCGA ACGACCGAGC GCAGCGAGTC
2761 AGTGAGCGAG GAAGCGGAAG AGCGCCTGAT GCGGTATTTT CTCCTTACGC ATCTGTGCGG
2821 TATTTCACAC CGCATATGGT GCACTCTCAG TACAATCTGC TCTGATGCCG CATAGTTAAG
2881 CCAGTATACA CTCCGCTATC GCTACGTGAC TGGGTCATGG CTGCGCCCCG ACACCCGCCA
2941 ACACCCGCTG ACGCGCCCTG ACGGGCTTGT CTGCTCCCGG CATCCGCTTA CAGACAAGCT
3001 GTGACCGTCT CCGGGAGCTG CATGTGTCAG AGGTTTTCAC CGTCATCACC GAAACGCGCG
3061 AGGCAGCAGA TCAATTCGCG CGCGAAGGCG AAGCGGCATG CATAATGTGC CTGTCAAATG
3121 GACGAAGCAG GGATTCTGCA AACCCTATGC TACTCCGTCA AGCCGTCAAT TGTCTGATTC
3181 GTTACCAATT ATGACAACTT GACGGCTACA TCATTCACTT TTTCTTCACA ACCGGCACGG
3241 AACTCGCTCG GGCTGGCCCC GGTGCATTTT TTAAATACCC GCGAGAAATA GAGTTGATCG
3301 TCAAAACCAA CATTGCGACC GACGGTGGCG ATAGGCATCC GGGTGGTGCT CAAAAGCAGC
3361 TTCGCCTGGC TGATACGTTG GTCCTCGCGC CAGCTTAAGA CGCTAATCCC TAACTGCTGG
3421 CGGAAAAGAT GTGACAGACG CGACGGCGAC AAGCAAACAT GCTGTGCGAC GCTGGCGATA
3481 TCAAAATTGC TGTCTGCCAG GTGATCGCTG ATGTACTGAC AAGCCTCGCG TACCCGATTA
3541 TCCATCGGTG GATGGAGCGA CTCGTTAATC GCTTCCATGC GCCGCAGTAA CAATTGCTCA
3601 AGCAGATTTA TCGCCAGCAG CTCCGAATAG CGCCCTTCCC CTTGCCCGGC GTTAATGATT
3661 TGCCCAAACA GGTCGCTGAA ATGCGGCTGG TGCGCTTCAT CCGGGCGAAA GAACCCCGTA
3721 TTGGCAAATA TTGACGGCCA GTTAAGCCAT TCATGCCAGT AGGCGCGCGG ACGAAAGTAA
3781 ACCCACTGGT GATACCATTC GCGAGCCTCC GGATGACGAC CGTAGTGATG AATCTCTCCT
3841 GGCGGGAACA GCAAAATATC ACCCGGTCGG CAAACAAATT CTCGTCCCTG ATTTTTCACC
3901 ACCCCCTGAC CGCGAATGGT GAGATTGAGA ATATAACCTT TCATTCCCAG CGGTCGGTCG
3961 ATAAAAAAAT CGAGATAACC GTTGGCCTCA ATCGGCGTTA AACCCGCCAC CAGATGGGCA
4021 TTAAACGAGT ATCCCGGCAG CAGGGGATCA TTTTGCGCTT CAGCCATACT TTTCATACTC
4081 CCGCCATTCA GAG
//
其他大肠杆菌表达载体:
