
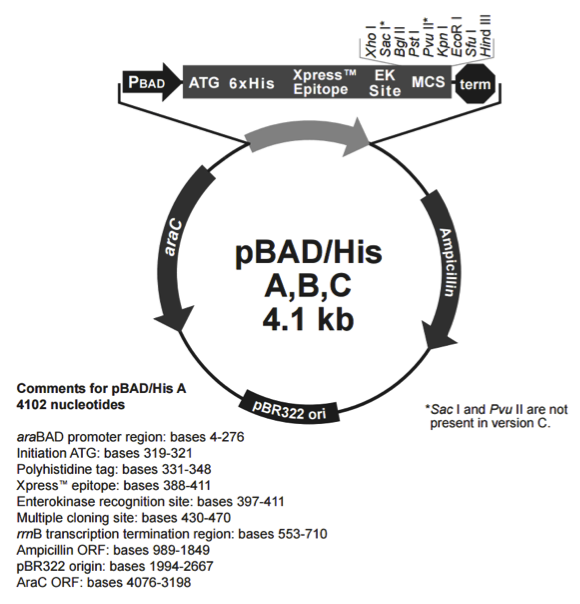
pBad/His C
pBad/His C
编号 | 载体名称 |
北京华越洋生物VECT4600 | pBad/His C |
pBadHis C载体基本信息
载体名称: | pBAD/His C |
质粒类型: | 大肠杆菌表达载体;诱导表达载体 |
高拷贝/低拷贝: | 低拷贝 |
克隆方法: | 限制性内切酶;多克隆位点 |
启动子: | araBAD |
载体大小: | 4100 bp |
5' 测序引物及序列: | pBAD Forward: 5′-ATGCCATAGCATTTTTATCC-3′ |
3' 测序引物及序列: | pBAD Reverse 5′-GATTTAATCTGTATCAGG-3′ |
载体标签: | 6x His Tag(N-端),Xpress Epitope(N-端),EK 切割位点 |
载体抗性: | 氨苄青霉素(Ampicillin) |
克隆菌株: | TOP10 |
表达菌株: | 推荐LMG194 |
备注: | pBAD/His C载体是阿拉伯糖调控载体; |
稳定性: | 稳表达 |
组成型/诱导型: | 诱导型(阿拉伯糖) |
病毒/非病毒: | 非病毒 |
pBadHis C载体质粒图谱和多克隆位点信息

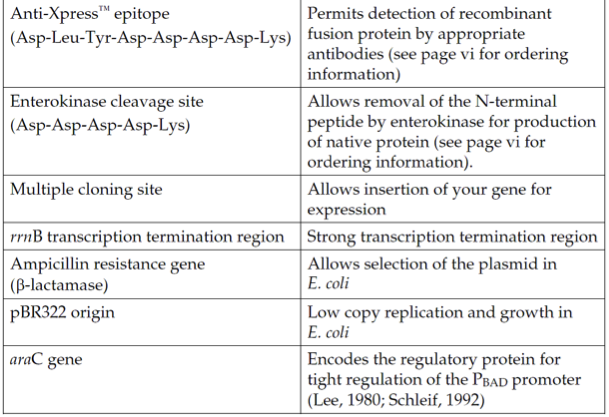
pBadHis C载体简介
pBAD/His和PBAD/Myc-His载体质粒是衍生于pBR322载体。载体设计用来在大肠杆菌中进行可调节,剂量依赖性的表达和纯化重组目的蛋白。使用大肠杆菌araBAD启动子(pBAD)增强了大肠杆菌重组蛋白可溶性表达的水平。pBAD/His和pBAD/Myc His载体上的调节蛋白AraC能够调控pBad启动子。
pBAD/His A,B,C 载体介绍
The pBAD/His Kit provides all of the necessary reagents to express your protein in a tightly regulated fashion. The vector pBAD/His allows you to express your protein with an N-terminal tag. The vector provides:
Þ The araBAD promoter for tightly regulated expression
Þ Translation initiation signals optimized for E. coliexpression
Þ N-terminal polyhistidine (6xHis) tag for purification with nickel-chelating resin and detection with an Anti-
HisG Antibody
Þ N-terminal Xpress epitope for detection and analysis with an Anti-Xpress Antibody
Þ Enterokinase cleavage site for removing the N-terminal tag following purification
Three vectors are provided (A, B, and C). Each has the N-terminal tag in a different reading frame relative to the multiple cloning site to simplify in-frame cloning of your gene.
The pBAD/His and pBAD/Myc-His plasmids are pBR322-derived expression vectors designed for regulated, dose-dependent recombinant protein expression and purification in E. coli. Optimum levels of soluble, recombinant protein are possible using the araBAD promoter (PBAD) from E. coli. The regulatory protein, AraC, is provided on the pBAD/His and pBAD/Myc-His vectors allowing regulation of PBAD.
L-阿拉伯糖调控表达
In the presence of L-arabinose, expression from PBAD is turned on while the absence of L-arabinose produces very low levels of transcription from PBAD (Lee, 1980; Lee et al., 1987). Uninduced levels are repressed even further by growth in the presence of glucose. Glucose reduces the levels of 3′,5′-cyclic AMP, thus lowering expression of the catabolite-repressed PBAD promoter (Miyada et al., 1984). By varying the concentration of L-arabinose, protein expression levels can be optimized to ensure maximum expression of soluble protein. In addition, the tight regulation of PBAD by AraC is useful for expression of potentially toxic or essential genes (Carson et al., 1991; Dalbey and Wickner, 1985; Guzman et al., 1992; Kuhn and Wickner, 1985; Russell et al., 1989; San Millan et al., 1989). For more information on the mechanism of expression and repression of the ara regulon, refer to Schleif, 1992.

pBadHis C载体序列
ORIGIN
1 AAGAAACCAA TTGTCCATAT TGCATCAGAC ATTGCCGTCA CTGCGTCTTT TACTGGCTCT
61 TCTCGCTAAC CAAACCGGTA ACCCCGCTTA TTAAAAGCAT TCTGTAACAA AGCGGGACCA
121 AAGCCATGAC AAAAACGCGT AACAAAAGTG TCTATAATCA CGGCAGAAAA GTCCACATTG
181 ATTATTTGCA CGGCGTCACA CTTTGCTATG CCATAGCATT TTTATCCATA AGATTAGCGG
241 ATCCTACCTG ACGCTTTTTA TCGCAACTCT CTACTGTTTC TCCATACCCG TTTTTTGGGC
301 TAACAGGAGG AATTAACCAT GGGGGGTTCT CATCATCATC ATCATCATGG TATGGCTAGC
361 ATGACTGGTG GACAGCAAAT GGGTCGGGAT CTGTACGACG ATGACGATAA GGATCGATGG
421 ATCCGACCTC GAGATCTGCA GATGGTACCA TATGGGAATT CGAAGCTTGG CTGTTTTGGC
481 GGATGAGAGA AGATTTTCAG CCTGATACAG ATTAAATCAG AACGCAGAAG CGGTCTGATA
541 AAACAGAATT TGCCTGGCGG CAGTAGCGCG GTGGTCCCAC CTGACCCCAT GCCGAACTCA
601 GAAGTGAAAC GCCGTAGCGC CGATGGTAGT GTGGGGTCTC CCCATGCGAG AGTAGGGAAC
661 TGCCAGGCAT CAAATAAAAC GAAAGGCTCA GTCGAAAGAC TGGGCCTTTC GTTTTATCTG
721 TTGTTTGTCG GTGAACGCTC TCCTGAGTAG GACAAATCCG CCGGGAGCGG ATTTGAACGT
781 TGCGAAGCAA CGGCCCGGAG GGTGGCGGGC AGGACGCCCG CCATAAACTG CCAGGCATCA
841 AATTAAGCAG AAGGCCATCC TGACGGATGG CCTTTTTGCG TTTCTACAAA CTCTTTTGTT
901 TATTTTTCTA AATACATTCA AATATGTATC CGCTCATGAG ACAATAACCC TGATAAATGC
961 TTCAATAATA TTGAAAAAGG AAGAGTATGA GTATTCAACA TTTCCGTGTC GCCCTTATTC
1021 CCTTTTTTGC GGCATTTTGC CTTCCTGTTT TTGCTCACCC AGAAACGCTG GTGAAAGTAA
1081 AAGATGCTGA AGATCAGTTG GGTGCACGAG TGGGTTACAT CGAACTGGAT CTCAACAGCG
1141 GTAAGATCCT TGAGAGTTTT CGCCCCGAAG AACGTTTTCC AATGATGAGC ACTTTTAAAG
1201 TTCTGCTATG TGGCGCGGTA TTATCCCGTG TTGACGCCGG GCAAGAGCAA CTCGGTCGCC
1261 GCATACACTA TTCTCAGAAT GACTTGGTTG AGTACTCACC AGTCACAGAA AAGCATCTTA
1321 CGGATGGCAT GACAGTAAGA GAATTATGCA GTGCTGCCAT AACCATGAGT GATAACACTG
1381 CGGCCAACTT ACTTCTGACA ACGATCGGAG GACCGAAGGA GCTAACCGCT TTTTTGCACA
1441 ACATGGGGGA TCATGTAACT CGCCTTGATC GTTGGGAACC GGAGCTGAAT GAAGCCATAC
1501 CAAACGACGA GCGTGACACC ACGATGCCTG TAGCAATGGC AACAACGTTG CGCAAACTAT
1561 TAACTGGCGA ACTACTTACT CTAGCTTCCC GGCAACAATT AATAGACTGG ATGGAGGCGG
1621 ATAAAGTTGC AGGACCACTT CTGCGCTCGG CCCTTCCGGC TGGCTGGTTT ATTGCTGATA
1681 AATCTGGAGC CGGTGAGCGT GGGTCTCGCG GTATCATTGC AGCACTGGGG CCAGATGGTA
1741 AGCCCTCCCG TATCGTAGTT ATCTACACGA CGGGGAGTCA GGCAACTATG GATGAACGAA
1801 ATAGACAGAT CGCTGAGATA GGTGCCTCAC TGATTAAGCA TTGGTAACTG TCAGACCAAG
1861 TTTACTCATA TATACTTTAG ATTGATTTAA AACTTCATTT TTAATTTAAA AGGATCTAGG
1921 TGAAGATCCT TTTTGATAAT CTCATGACCA AAATCCCTTA ACGTGAGTTT TCGTTCCACT
1981 GAGCGTCAGA CCCCGTAGAA AAGATCAAAG GATCTTCTTG AGATCCTTTT TTTCTGCGCG
2041 TAATCTGCTG CTTGCAAACA AAAAAACCAC CGCTACCAGC GGTGGTTTGT TTGCCGGATC
2101 AAGAGCTACC AACTCTTTTT CCGAAGGTAA CTGGCTTCAG CAGAGCGCAG ATACCAAATA
2161 CTGTCCTTCT AGTGTAGCCG TAGTTAGGCC ACCACTTCAA GAACTCTGTA GCACCGCCTA
2221 CATACCTCGC TCTGCTAATC CTGTTACCAG TGGCTGCTGC CAGTGGCGAT AAGTCGTGTC
2281 TTACCGGGTT GGACTCAAGA CGATAGTTAC CGGATAAGGC GCAGCGGTCG GGCTGAACGG
2341 GGGGTTCGTG CACACAGCCC AGCTTGGAGC GAACGACCTA CACCGAACTG AGATACCTAC
2401 AGCGTGAGCT ATGAGAAAGC GCCACGCTTC CCGAAGGGAG AAAGGCGGAC AGGTATCCGG
2461 TAAGCGGCAG GGTCGGAACA GGAGAGCGCA CGAGGGAGCT TCCAGGGGGA AACGCCTGGT
2521 ATCTTTATAG TCCTGTCGGG TTTCGCCACC TCTGACTTGA GCGTCGATTT TTGTGATGCT
2581 CGTCAGGGGG GCGGAGCCTA TGGAAAAACG CCAGCAACGC GGCCTTTTTA CGGTTCCTGG
2641 CCTTTTGCTG GCCTTTTGCT CACATGTTCT TTCCTGCGTT ATCCCCTGAT TCTGTGGATA
2701 ACCGTATTAC CGCCTTTGAG TGAGCTGATA CCGCTCGCCG CAGCCGAACG ACCGAGCGCA
2761 GCGAGTCAGT GAGCGAGGAA GCGGAAGAGC GCCTGATGCG GTATTTTCTC CTTACGCATC
2821 TGTGCGGTAT TTCACACCGC ATATGGTGCA CTCTCAGTAC AATCTGCTCT GATGCCGCAT
2881 AGTTAAGCCA GTATACACTC CGCTATCGCT ACGTGACTGG GTCATGGCTG CGCCCCGACA
2941 CCCGCCAACA CCCGCTGACG CGCCCTGACG GGCTTGTCTG CTCCCGGCAT CCGCTTACAG
3001 ACAAGCTGTG ACCGTCTCCG GGAGCTGCAT GTGTCAGAGG TTTTCACCGT CATCACCGAA
3061 ACGCGCGAGG CAGCAGATCA ATTCGCGCGC GAAGGCGAAG CGGCATGCAT AATGTGCCTG
3121 TCAAATGGAC GAAGCAGGGA TTCTGCAAAC CCTATGCTAC TCCGTCAAGC CGTCAATTGT
3181 CTGATTCGTT ACCAATTATG ACAACTTGAC GGCTACATCA TTCACTTTTT CTTCACAACC
3241 GGCACGGAAC TCGCTCGGGC TGGCCCCGGT GCATTTTTTA AATACCCGCG AGAAATAGAG
3301 TTGATCGTCA AAACCAACAT TGCGACCGAC GGTGGCGATA GGCATCCGGG TGGTGCTCAA
3361 AAGCAGCTTC GCCTGGCTGA TACGTTGGTC CTCGCGCCAG CTTAAGACGC TAATCCCTAA
3421 CTGCTGGCGG AAAAGATGTG ACAGACGCGA CGGCGACAAG CAAACATGCT GTGCGACGCT
3481 GGCGATATCA AAATTGCTGT CTGCCAGGTG ATCGCTGATG TACTGACAAG CCTCGCGTAC
3541 CCGATTATCC ATCGGTGGAT GGAGCGACTC GTTAATCGCT TCCATGCGCC GCAGTAACAA
3601 TTGCTCAAGC AGATTTATCG CCAGCAGCTC CGAATAGCGC CCTTCCCCTT GCCCGGCGTT
3661 AATGATTTGC CCAAACAGGT CGCTGAAATG CGGCTGGTGC GCTTCATCCG GGCGAAAGAA
3721 CCCCGTATTG GCAAATATTG ACGGCCAGTT AAGCCATTCA TGCCAGTAGG CGCGCGGACG
3781 AAAGTAAACC CACTGGTGAT ACCATTCGCG AGCCTCCGGA TGACGACCGT AGTGATGAAT
3841 CTCTCCTGGC GGGAACAGCA AAATATCACC CGGTCGGCAA ACAAATTCTC GTCCCTGATT
3901 TTTCACCACC CCCTGACCGC GAATGGTGAG ATTGAGAATA TAACCTTTCA TTCCCAGCGG
3961 TCGGTCGATA AAAAAATCGA GATAACCGTT GGCCTCAATC GGCGTTAAAC CCGCCACCAG
4021 ATGGGCATTA AACGAGTATC CCGGCAGCAG GGGATCATTT TGCGCTTCAG CCATACTTTT
4081 CATACTCCCG CCATTCAGAG
//
pBad/His C其他大肠杆菌表达载体:
