

pBad/Myc-His B
pBad/Myc-His B
编号 | 载体名称 |
北京华越洋生物VECT4060 | pBad/Myc-His B |
pBadMyc-His B载体基本信息
载体名称: | pBAD/Myc-His B |
质粒类型: | 大肠杆菌表达载体;诱导表达载体 |
高拷贝/低拷贝: | 低拷贝 |
克隆方法: | 限制性内切酶;多克隆位点 |
启动子: | araBAD |
载体大小: | 4092 bp |
5' 测序引物及序列: | pBAD Forward: 5′-ATGCCATAGCATTTTTATCC-3′ |
3' 测序引物及序列: | pBAD Reverse 5′-GATTTAATCTGTATCAGG-3′ |
载体标签: | 6x His Tag(C-端),c-Myc Epitope(C-端) |
载体抗性: | 氨苄青霉素(Ampicillin) |
克隆菌株: | TOP10 |
表达菌株: | 推荐LMG194 |
备注: | pBAD/Myc-His B载体是阿拉伯糖调控载体;在无葡萄糖的培养基中,阿拉伯糖正向调控目 的基因的表达;通过调节阿拉伯糖的浓度水平来优化目的蛋白的可溶性表达。 |
稳定性: | 稳表达 |
组成型/诱导型: | 诱导型(阿拉伯糖) |
病毒/非病毒: | 非病毒 |
pBadMyc-His B载体质粒图谱和多克隆位点信息


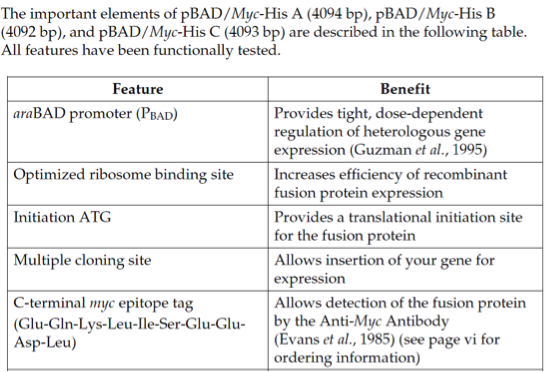

pBadMyc-His B载体简介
pBAD/His和PBAD/Myc-His载体质粒是衍生于pBR322载体。载体设计用来在大肠杆菌中进行可调节,剂量依赖性的表达和纯化重组目的蛋 白。使用大肠杆菌araBAD启动子(pBAD)增强了大肠杆菌重组蛋白可溶性表达的水平。pBAD/His和pBAD/Myc His载体上的调节蛋白AraC能够调控pBad启动子。
pBAD/Myc-His A,B,C 载体简介
The pBAD/His and pBAD/Myc-His plasmids are pBR322-derived expression vectors designed for regulated, dose-dependent recombinant protein expression and purification in E. coli. Optimum levels of soluble, recombinant protein are possible using the araBAD promoter (PBAD) from E. coli. The regulatory protein, AraC, is provided on the pBAD/His and pBAD/Myc-His vectors allowing regulation of PBAD.
The pBAD/Myc-His Kit provides all of the necessary reagents to express your protein in a tightly regulated fashion. The pBAD/Myc-His vector expresses native proteins or fusion proteins with a C-terminal tag. The vector provides:
Þ The araBAD promoter for tightly regulated expression
Þ Translation initiation signals optimized for E. coliexpression
Þ C-terminal polyhistidine (6xHis) tag for purification with nickel-chelating resin or detection with an Anti-His(C-term) Antibody
Þ C-terminal c-myc epitope for detection and analysis with an Anti-myc Antibody
Þ Three vectors are provided (A, B, and C). Each has the C-terminal tag in a different reading frame relative to the multiple cloning site to simplify in-frame cloning of your gene.
L-阿拉伯糖调控表达
In the presence of L-arabinose, expression from PBAD is turned on while the absence of L-arabinose produces very low levels of transcription from PBAD (Lee, 1980; Lee et al., 1987). Uninduced levels are repressed even further by growth in the presence of glucose. Glucose reduces the levels of 3′,5′-cyclic AMP, thus lowering expression of the catabolite-repressed PBAD promoter (Miyada et al., 1984). By varying the concentration of L-arabinose, protein expression levels can be optimized to ensure maximum expression of soluble protein. In addition, the tight regulation of PBAD by AraC is useful for expression of potentially toxic or essential genes (Carson et al., 1991; Dalbey and Wickner, 1985; Guzman et al., 1992; Kuhn and Wickner, 1985; Russell et al., 1989; San Millan et al., 1989). For more information on the mechanism of expression and repression of the ara regulon, refer to Schleif, 1992.
pBadMyc-His B载体序列
ORIGIN
1 AAGAAACCAA TTGTCCATAT TGCATCAGAC ATTGCCGTCA CTGCGTCTTT TACTGGCTCT
61 TCTCGCTAAC CAAACCGGTA ACCCCGCTTA TTAAAAGCAT TCTGTAACAA AGCGGGACCA
121 AAGCCATGAC AAAAACGCGT AACAAAAGTG TCTATAATCA CGGCAGAAAA GTCCACATTG
181 ATTATTTGCA CGGCGTCACA CTTTGCTATG CCATAGCATT TTTATCCATA AGATTAGCGG
241 ATCCTACCTG ACGCTTTTTA TCGCAACTCT CTACTGTTTC TCCATACCCG TTTTTTGGGC
301 TAACAGGAGG AATTAACCAT GGATCCGAGC TCGAGATCTG CAGCTGGTAC CATATGGGAA
361 TTCGAAGCTT TCTAGAACAA AAACTCATCT CAGAAGAGGA TCTGAATAGC GCCGTCGACC
421 ATCATCATCA TCATCATTGA GTTTAAACGG TCTCCAGCTT GGCTGTTTTG GCGGATGAGA
481 GAAGATTTTC AGCCTGATAC AGATTAAATC AGAACGCAGA AGCGGTCTGA TAAAACAGAA
541 TTTGCCTGGC GGCAGTAGCG CGGTGGTCCC ACCTGACCCC ATGCCGAACT CAGAAGTGAA
601 ACGCCGTAGC GCCGATGGTA GTGTGGGGTC TCCCCATGCG AGAGTAGGGA ACTGCCAGGC
661 ATCAAATAAA ACGAAAGGCT CAGTCGAAAG ACTGGGCCTT TCGTTTTATC TGTTGTTTGT
721 CGGTGAACGC TCTCCTGAGT AGGACAAATC CGCCGGGAGC GGATTTGAAC GTTGCGAAGC
781 AACGGCCCGG AGGGTGGCGG GCAGGACGCC CGCCATAAAC TGCCAGGCAT CAAATTAAGC
841 AGAAGGCCAT CCTGACGGAT GGCCTTTTTG CGTTTCTACA AACTCTTTTG TTTATTTTTC
901 TAAATACATT CAAATATGTA TCCGCTCATG AGACAATAAC CCTGATAAAT GCTTCAATAA
961 TATTGAAAAA GGAAGAGTAT GAGTATTCAA CATTTCCGTG TCGCCCTTAT TCCCTTTTTT
1021 GCGGCATTTT GCCTTCCTGT TTTTGCTCAC CCAGAAACGC TGGTGAAAGT AAAAGATGCT
1081 GAAGATCAGT TGGGTGCACG AGTGGGTTAC ATCGAACTGG ATCTCAACAG CGGTAAGATC
1141 CTTGAGAGTT TTCGCCCCGA AGAACGTTTT CCAATGATGA GCACTTTTAA AGTTCTGCTA
1201 TGTGGCGCGG TATTATCCCG TGTTGACGCC GGGCAAGAGC AACTCGGTCG CCGCATACAC
1261 TATTCTCAGA ATGACTTGGT TGAGTACTCA CCAGTCACAG AAAAGCATCT TACGGATGGC
1321 ATGACAGTAA GAGAATTATG CAGTGCTGCC ATAACCATGA GTGATAACAC TGCGGCCAAC
1381 TTACTTCTGA CAACGATCGG AGGACCGAAG GAGCTAACCG CTTTTTTGCA CAACATGGGG
1441 GATCATGTAA CTCGCCTTGA TCGTTGGGAA CCGGAGCTGA ATGAAGCCAT ACCAAACGAC
1501 GAGCGTGACA CCACGATGCC TGTAGCAATG GCAACAACGT TGCGCAAACT ATTAACTGGC
1561 GAACTACTTA CTCTAGCTTC CCGGCAACAA TTAATAGACT GGATGGAGGC GGATAAAGTT
1621 GCAGGACCAC TTCTGCGCTC GGCCCTTCCG GCTGGCTGGT TTATTGCTGA TAAATCTGGA
1681 GCCGGTGAGC GTGGGTCTCG CGGTATCATT GCAGCACTGG GGCCAGATGG TAAGCCCTCC
1741 CGTATCGTAG TTATCTACAC GACGGGGAGT CAGGCAACTA TGGATGAACG AAATAGACAG
1801 ATCGCTGAGA TAGGTGCCTC ACTGATTAAG CATTGGTAAC TGTCAGACCA AGTTTACTCA
1861 TATATACTTT AGATTGATTT AAAACTTCAT TTTTAATTTA AAAGGATCTA GGTGAAGATC
1921 CTTTTTGATA ATCTCATGAC CAAAATCCCT TAACGTGAGT TTTCGTTCCA CTGAGCGTCA
1981 GACCCCGTAG AAAAGATCAA AGGATCTTCT TGAGATCCTT TTTTTCTGCG CGTAATCTGC
2041 TGCTTGCAAA CAAAAAAACC ACCGCTACCA GCGGTGGTTT GTTTGCCGGA TCAAGAGCTA
2101 CCAACTCTTT TTCCGAAGGT AACTGGCTTC AGCAGAGCGC AGATACCAAA TACTGTCCTT
2161 CTAGTGTAGC CGTAGTTAGG CCACCACTTC AAGAACTCTG TAGCACCGCC TACATACCTC
2221 GCTCTGCTAA TCCTGTTACC AGTGGCTGCT GCCAGTGGCG ATAAGTCGTG TCTTACCGGG
2281 TTGGACTCAA GACGATAGTT ACCGGATAAG GCGCAGCGGT CGGGCTGAAC GGGGGGTTCG
2341 TGCACACAGC CCAGCTTGGA GCGAACGACC TACACCGAAC TGAGATACCT ACAGCGTGAG
2401 CTATGAGAAA GCGCCACGCT TCCCGAAGGG AGAAAGGCGG ACAGGTATCC GGTAAGCGGC
2461 AGGGTCGGAA CAGGAGAGCG CACGAGGGAG CTTCCAGGGG GAAACGCCTG GTATCTTTAT
2521 AGTCCTGTCG GGTTTCGCCA CCTCTGACTT GAGCGTCGAT TTTTGTGATG CTCGTCAGGG
2581 GGGCGGAGCC TATGGAAAAA CGCCAGCAAC GCGGCCTTTT TACGGTTCCT GGCCTTTTGC
2641 TGGCCTTTTG CTCACATGTT CTTTCCTGCG TTATCCCCTG ATTCTGTGGA TAACCGTATT
2701 ACCGCCTTTG AGTGAGCTGA TACCGCTCGC CGCAGCCGAA CGACCGAGCG CAGCGAGTCA
2761 GTGAGCGAGG AAGCGGAAGA GCGCCTGATG CGGTATTTTC TCCTTACGCA TCTGTGCGGT
2821 ATTTCACACC GCATATGGTG CACTCTCAGT ACAATCTGCT CTGATGCCGC ATAGTTAAGC
2881 CAGTATACAC TCCGCTATCG CTACGTGACT GGGTCATGGC TGCGCCCCGA CACCCGCCAA
2941 CACCCGCTGA CGCGCCCTGA CGGGCTTGTC TGCTCCCGGC ATCCGCTTAC AGACAAGCTG
3001 TGACCGTCTC CGGGAGCTGC ATGTGTCAGA GGTTTTCACC GTCATCACCG AAACGCGCGA
3061 GGCAGCAGAT CAATTCGCGC GCGAAGGCGA AGCGGCATGC ATAATGTGCC TGTCAAATGG
3121 ACGAAGCAGG GATTCTGCAA ACCCTATGCT ACTCCGTCAA GCCGTCAATT GTCTGATTCG
3181 TTACCAATTA TGACAACTTG ACGGCTACAT CATTCACTTT TTCTTCACAA CCGGCACGGA
3241 ACTCGCTCGG GCTGGCCCCG GTGCATTTTT TAAATACCCG CGAGAAATAG AGTTGATCGT
3301 CAAAACCAAC ATTGCGACCG ACGGTGGCGA TAGGCATCCG GGTGGTGCTC AAAAGCAGCT
3361 TCGCCTGGCT GATACGTTGG TCCTCGCGCC AGCTTAAGAC GCTAATCCCT AACTGCTGGC
3421 GGAAAAGATG TGACAGACGC GACGGCGACA AGCAAACATG CTGTGCGACG CTGGCGATAT
3481 CAAAATTGCT GTCTGCCAGG TGATCGCTGA TGTACTGACA AGCCTCGCGT ACCCGATTAT
3541 CCATCGGTGG ATGGAGCGAC TCGTTAATCG CTTCCATGCG CCGCAGTAAC AATTGCTCAA
3601 GCAGATTTAT CGCCAGCAGC TCCGAATAGC GCCCTTCCCC TTGCCCGGCG TTAATGATTT
3661 GCCCAAACAG GTCGCTGAAA TGCGGCTGGT GCGCTTCATC CGGGCGAAAG AACCCCGTAT
3721 TGGCAAATAT TGACGGCCAG TTAAGCCATT CATGCCAGTA GGCGCGCGGA CGAAAGTAAA
3781 CCCACTGGTG ATACCATTCG CGAGCCTCCG GATGACGACC GTAGTGATGA ATCTCTCCTG
3841 GCGGGAACAG CAAAATATCA CCCGGTCGGC AAACAAATTC TCGTCCCTGA TTTTTCACCA
3901 CCCCCTGACC GCGAATGGTG AGATTGAGAA TATAACCTTT CATTCCCAGC GGTCGGTCGA
3961 TAAAAAAATC GAGATAACCG TTGGCCTCAA TCGGCGTTAA ACCCGCCACC AGATGGGCAT
4021 TAAACGAGTA TCCCGGCAGC AGGGGATCAT TTTGCGCTTC AGCCATACTT TTCATACTCC
4081 CGCCATTCAG AG
//
pBad/Myc-His B其他大肠杆菌表达载体:
